








概述
对于家禽生产行业,啄羽是一种严重问题。啄羽行为会造成巨大的经济损失,也会给受影响的禽类带来痛苦,尤其是在禁止断喙的国家。 尽管进行了深入研究,但这种破坏性行为的原因仍未完全明确。大多数解决问题的方法将重点放在营养和管理方面。对于啄羽行为,所有措施只能起到缓解作用或没有效果。遗传学研究表明,对于这种破坏性行为,充分的遗传变异能够实现遗传选育。
然而,啄羽是一种复杂行为,如果需要通过育种计划成功体现这一性状,还需要了解更多关于动机遗传背景的详细信息。主流假说认为啄羽是定向错误的觅食行为,但也可能涉及如食羽癖、攻击性行为、恐惧和常规自发活动等其他动机。
通过两个F2杂交品系的900多只禽类(根据啄羽行为表现强弱进行选育)研究了上述行为之间的相互关系。使用标准统计模型对于这些性状之间的遗传性以及遗传和表型相关性进行了估测。此外,同时应用了结构方程模型(SEM)以估测啄羽与其他性状之间的因果关系。
遗传相关性和作为因果联系参数的λ系数表明,食羽癖和啄羽行为之间具有很强的因果关系。这证实了食羽癖是啄羽主要原因的假说。 攻击性行为和常规自发活动也对啄羽产生很大的因果影响。相反,旷场行为(恐惧)和觅食对于啄羽行为并未产生明显影响。
关键词
蛋鸡、啄羽、攻击性行为、恐惧、活动、探索、遗传学共同作者
J.B. Kjaer(FLI,策勒)、V. Lutz(霍恩海姆大学)、M. Grashorn(霍恩海姆大学)、W. Bessei(霍恩海姆大学)
引言
对于工业化国家的家禽生产行业, 动物福利已成为一个重要的问题。有人认为集约化管理体系和生产性状选育会损害动物福利。例如,快速生长肉鸡的软腿病、腹水和猝死(Bessei和Gerken,2006年)以及蛋鸡的骨折(Moinard等,1998年),都说明业绩和动物福利相关性状之间存在对立关系。基因组工具的开发有助于开展这样的讨论,而这种工具应用于商业育种将进一步提升业绩。过去,高业绩一直被视为动物福利的间接标准。然而,人们日益意识到,针对生长率或产蛋率的选育可能会导致动物福利问题,尤其是“精准动物育种”框架(Flint和Woolliams,2007年)和SEFABAR项目(“欧洲农场动物的可持续育种和繁殖”,SEFABAR,2003年),体现了科学家对于农场动物福利的责任感。 经济标准和动物福利标准并不一定是对立的。例如,对于家禽类,破坏性的啄羽行为和同类相残行为是严重的动物福利问题,可能对经济业绩产生重要影响。如果出于动物福利原因禁止断喙,这个问题将变得更加重要。最近针对未断喙鸡的实验表明,管理流程可减少啄羽现象,但不能杜绝啄羽。由于啄羽具有很强的遗传因素,业界正在寻找将这种行为纳入选育计划的方法。了解啄羽的原因是解决问题的前提条件。 下面,我们将利用Gianola和Soerensen(2004年)提出的结构方程模型,对于啄羽行为以及啄羽行为和选定行为标准之间的因果关系进行定量遗传分析。
信息状况和实验方法
破坏性啄羽是一种复杂的行为。尽管在过去几十年中进行了大量研究,但其根本原因仍不得而知。 不过,关于其动机存在各种假说。主流理论认为啄羽与喂食和饲料搜寻(觅食)有关(Blokhuis,1986年;Huber-Eicher和Wechsler,1998年)。觅食理论认为觅食行为会转向群体同伴的羽毛。但是,在多项研究中,觅食理论都无法解释啄羽现象(Hocking等,2004年;Newberry等,2007年)。Bessei和Kjaer(2015年)认为食羽癖是主要的潜在动机。目前纳入讨论范围的啄羽动机还包括攻击性行为(Bessei等,2013年)、恐惧(Rodenburg等,2010年)和自发性活动(Kjaer,2009年)。啄羽行为取决于沙浴的假说(Vestergaard和Lisborg,1993年)在后来的实验中没有得到证实,因此不再考虑。关于啄羽行为与其他行为之间的上述关联性假设,主要基于群体平均值对比和常规相关性估测,而常规相关性估测并不能揭示因果关系。 然而,同样有一些方法能够估测性状之间的因果效应(Gianola和Soerensen,2004年),例如结构方程模型(SEM)。在一个来自两个根据啄羽行为表现强弱进行选育的品系的大型F2杂交种群(大于900只鸡)中,观察到了啄羽(FP)、食羽癖(FE)、攻击性啄羽(AP)、旷场行为(恐惧)(OFA)、常规自发活动(GLA)和觅食(FOR)(图1)。采用标准多性状模型(SMS)和结构方程模型(SEM)对于遗传参数进行了估算,以揭示上述行为之间的因果关系。如果应用SEM,需要了解假说的精确先验定义。因此,我们采用了每组三种行为的测试方法。只选择存在假定的因果关系的行为进行研究。

啄羽、食羽癖和攻击性啄羽
先前研究表明,啄羽和食羽癖密切相关。研究发现,对同伴有严重啄羽倾向的禽类在单独饲养时,会吃掉塑料盘中的更多羽毛(McKeegan和Savory,2001年)。 在选择实验中,啄羽表现较强的蛋鸡更喜欢羽毛而不是刨花,而且从塑料盘中吃掉的羽毛比啄羽表现较弱的蛋鸡更多(Harlander-Matauschek和Häusler,2009年)。根据这些结果,我们认为食羽癖是影响啄羽的主要动机。人们普遍认为,攻击性啄羽和啄羽在表型模式上明显不同,其动机也不同(Savory,1995年;Bilcik和Keeling,1999年)。定量遗传研究和选择实验证明攻击性行为具有遗传性(Craig等,1965年)。Bennewitz等(2014年)和Grams等(2014年)对结果进行了详细报告。根据统计模型的不同,FP的遗传率估算值在0.11和0.20之间。 FE 的遗传率要高得多(0.36至0.57)。对于FE和AP,FP具有非常高的遗传相关性。所有表型相关性均为正相关,但水平低于遗传相关性。根据以往信息,我们认为FE会对FP和AP产生影响,而AP会影响FP。FE对于FP的估测因果效应系数λ(FP、FE)为5.94(图2)。关于食羽癖对于攻击性啄羽的影响系数λ(AP、FE)和攻击性啄羽对于啄羽的影响系数λ(FP、AP),λ-值均为正值(0.11和0.23),但大大低于食羽癖对于啄羽的影响。结果证实了我们的假设,即FE是FP的主要动机。FE对于AP的影响也很小。啄羽和攻击性啄羽之间的高度遗传相关性以及正值λ系数(FP、AP)表明,攻击性行为的影响在过去估计不足。在运动模式和潜在动机上,虽然攻击性行为与啄羽有所不同,但攻击性行为可能会强化FP的表达。
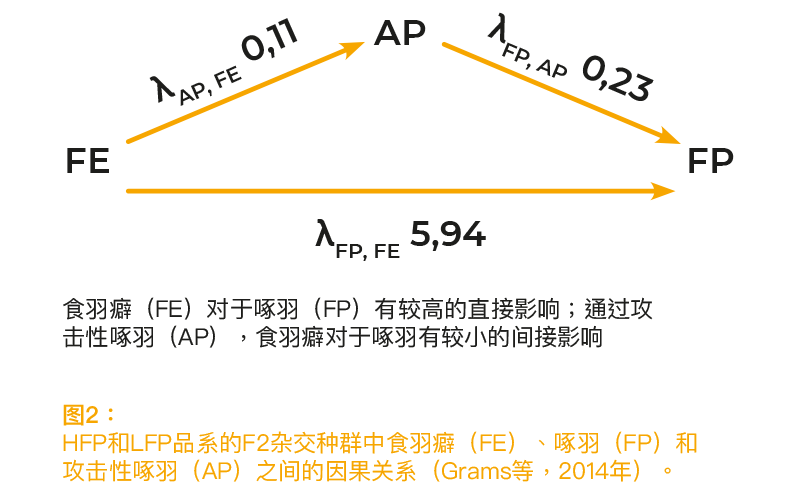
啄羽、食羽癖和常规自发活动
自发活动通常是由多种不同的动机激发的,例如探索、攻击性行为、产蛋和飞行。恐惧会抑制自发活动,具体情况请参考下文。 在实际条件下,不可能确定潜在的动机。动物的自发活动水平存在个体差异,这与特定的动机无关。这种“常规自发活动”(GLA)可认定为基因决定的自发性活动的结果。McClearn(1961年)(使用小鼠)以及Jezierski和Bessei(1978年)报告了GLA的遗传估算值。Kjaer(2009年)使用相同的记录体系,分别选择了GLA表现强和弱的鸡品系。遗传系数的估算值为0.20至0.30(个人通信)。作者认为,啄羽可能是多动症的结果。生理学研究支持了这一假说,研究表明,啄羽和过度活跃都受到多巴胺系统的影响(Kjaer等,2004年;van Hierden等,2005年)。使用了上述F2杂交小母鸡的自发活动长时间记录,以验证这一假说(Lutz等,2016年)。 18周龄时,使用RFID技术连续9天记录自发活动的情况。GLA的遗传率为0.29,与FP之间的表型和遗传相关性均呈正相关(0.16和0.47)。关于性状之间的因果关系,我们认为GLA会影响FP,而FE会影响FP和GLA。通过SEM获得的结果如图3所示。结构系数显示,GLA对于FP有因果影响(λFP、GLA = 1.20),FE对于FP有因果影响(λ = 5.71)。FE对于GLA的影响较小。结果证实了GLA会影响FP的假说。然而,FE(作为主要动机)对于FP的影响并不是通过GLA介导的。GLA对于FP的影响明显小于FE对于FP的影响。这两种影响似乎是相互独立的。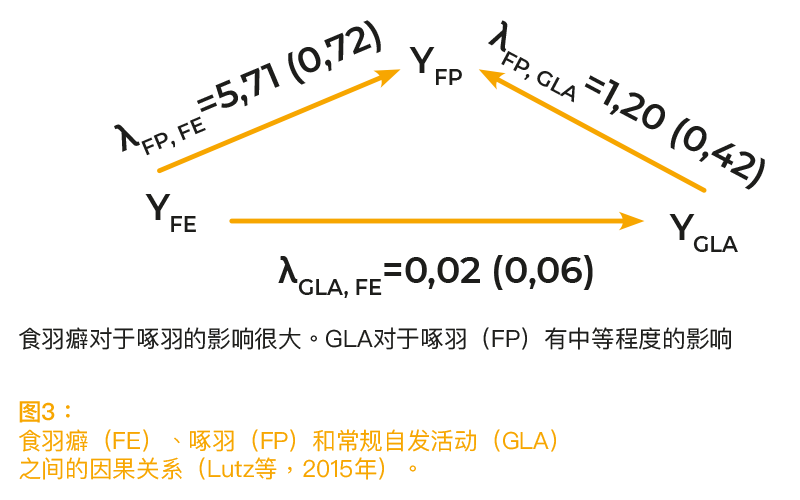
啄羽、觅食和恐惧
对于野生动物的生存策略,觅食(FOR)和恐惧是非常重要的因素。高度恐惧会抑制动物对于未知饲料资源的探索和利用。然而,在捕食者的高压下,恐惧是生存的必要条件。 因此,在自然条件下观察到的觅食活动可认定为探索和恐惧之间的折衷方案。最近观察表明,这种机制在自由放养的家禽中仍然存在。在惊吓瘫痪测试中表现出高度恐惧的母鸡,在自由放养环境中花费的时间较少(Hartcher等,2016年)。多种测试均能对动物的恐惧感进行评估。对于实验室动物和家禽,旷场测试是常用的测试方法。使动物暴露在旷场环境中,会引起动物的初始恐惧反应,即一动不动,随后会出现自发活动。高旷场行为(OFA)表明恐惧程度较低,可认定为探索性活动(Jones,1989年)。因矿场测试对探索行为产生影响,旷场测试适用于本研究。此外,据报道,若雏鸡OFA发生率较高,那么啄羽的发生率很可能较低(Rodenburg等,2004年)。 因此,对于啄羽表现强和表现弱的选育品系,我们使用SEM对于F2杂交种群的FP、OFA和FOR数据进行了分析。在7和8日龄时,以3分钟测试中的步数记录衡量OFA。FOR指的是:3周龄和4周龄时,在20分钟的观察时间内,在家庭围栏内行走和啄垃圾的时间。假设OFA和FOR影响FP,而FOR会影响OFA。FOR的遗传率为零。因此,无法估测这一性状的遗传相关性。OFA的遗传率为 0.21。 FP、OFA和FOR之间的表型相关性以及FP和OFA之间的遗传相关性均接近于零(Grams等,2014年)。所有λ系数均接近于零(图4)。结果表明,高估了觅食和恐惧对啄羽的影响。与主流假说相反,FOR和OFA均未显示出对于FP的相关因果效应。
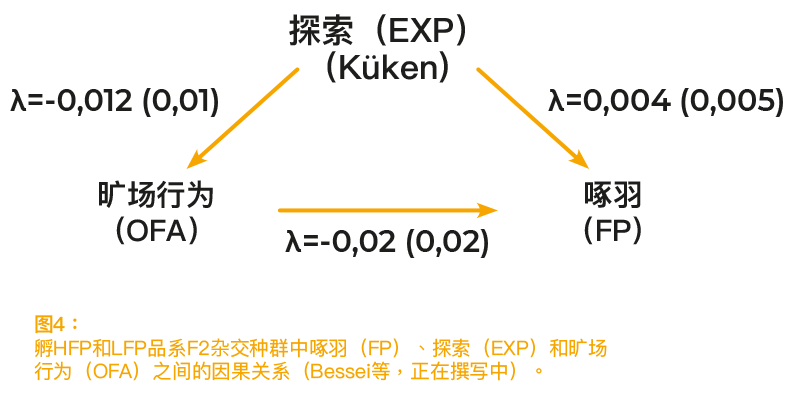
参考文献
BENNEWITZ,J.、BÖGELEIN, S.、STRATZ, P.、RODEHUTSCORD, M.、PIEPHO, H.P.、KJAER, J.B.和BESSEI, W.(2014年)使用广义线性混合模型计算蛋鸡大型F2杂交种群中关于啄羽和攻击性行为的遗传参数。《家禽科学》第93期:810-817。BESSEI, W.(2013年)啄羽表现强弱选育对于蛋鸡攻击性相关行为的影响。Archiv für Geflügelkunde 77:10; 10-14; 14。
BESSEI, W.和KJAER, J.B.(2015年)蛋鸡啄羽 – 研究现状与启示。26: 方法澳大利亚家禽科学研讨会,澳大利亚悉尼,2月9至12日。2015:214-221。
BILCIK, B.和KEELING, L.J.(1999年)与蛋鸡啄羽和攻击性行为有关的羽毛状况变化。《英国家禽科学》第40期:444-451。
BLOKHUIS, H.J.(1986年)家禽的啄羽:与啄地之间的关系。《应用动物行为科学》第16期:63-?。
CRAIG, J.V.、ORTMAN, L.L.和GUHL, A.M.(1965年)鸡群社会支配能力的基因选育。《家禽科学》13: 114-131。
DAWKINS, M.S.和LAYTON, R.(2012年)旨在提升动物福利的育种:肉鸡及其亲代的遗传目标。《动物福利》第21期:147-155。
ELLEN, E.D.、VISSCHER, J.、VAN ARENDONK, J.A.M.和BIJMA, P.(2008年)蛋鸡的存活率:三个纯种蛋鸡品系中直接效应和关联效应的遗传参数。87: 233-239。
GERKEN, M.和BESSEI, W.(2006年)肉用家禽的动物福利。WPSA – 第十二届欧洲家禽大会摘要手册,2006年9月10至14日,维罗纳:582-?。
GIANOLA, D.和SOERENSEN, D.(2004年)描述表型之间的联合关系和递归关系的定量遗传模型。167: 1407-1424。
Iba, A.M.和Berchieri, A.(1995年)关于使用甲酸-丙酸混合物(Bio-Add™)控制肉鸡沙门氏菌感染(基于实验)的研究。《禽类病理学》第24期,第303至311页。
GRAMS, V.、BESSEI, W.、PIEPHO, H.-.和BENNEWITZ, J.(2014年)通过泊松模型和线性模型确定关于蛋鸡啄羽和攻击性行为的遗传参数。方法第十届世界畜牧生产遗传学应用大会,2014年8月17至22日,加拿大温哥华。
GRAMS, V.、BÖGELEIN, S.、GRASHORN, M.A.、BESSEI, W.和BENNEWITZ, J.(2015年)关于蛋鸡恐惧和啄羽的定量遗传分析。《行为遗传学》第45期:228-235。
HARLANDER-MATAUSCHEK, A.和HÄUSLER, K.(2009年)了解蛋鸡食羽癖行为117:35-?。
HARTCHER, K.M.、HICKEY, K.A.、HEMSWORTH, P.H.、CRONIN, G.M.、WILKINSON, S.J.和SINGH, M.(2016年)自由放养蛋鸡的活动范围、恐惧和羽毛损伤(通过射频识别技术监测)之间的关系。《动物》第10期:847-853。
HOCKING, P.M.、CHANNING, C.E.、ROBERTSON, G.W.、EDMOND, A.和JONES, R.B.(2004年)家禽福利相关性状的品种间遗传变异。《应用动物行为科学》第89期:85-105。
JONES,R.B.(1989年)鸟类旷场研究及对于环境新奇性的相关影响。《心理学记录》第39期:397-420。
KJAER, J.B.(2009年)家禽啄羽行为与自发活动水平之间具有遗传相关性:啄羽多动症模型的意义。《行为遗传学》第39期:564-570。
LUTZ, V.、KJAER, J.B.、IFFLAND, H.、RODEHUTSCORD, M.、BESSEI, W.和BENNEWITZ, J.(2016年)利用结构方程模型对于蛋鸡啄羽、食羽癖和常规自发活动之间的因果关系进行定性遗传分析。《家禽科学》第95期:1757-1763。
MCKEEGAN, D.E.F.和SAVORY, C.J.(2001年)啄羽倾向性不同的单独笼养母鸡的食羽癖。期刊?73: 131-140。
MCKEEGAN, D.E.F.、SAVORY, C.J.、MACLEOD, M.G.和MITCHELL, M.A.(1999年)年轻蛋鸡的啄伤行为是否会受到食源性蛋白来源的影响?《英国家禽科学》第40期:14-15。
MOINARD, C.、MORISSE, J.P.和FAURE, J.M.(1998年)笼子面积、高度和栖木对于蛋鸡羽毛状况、骨折和死亡率的影响。《英国家禽科学》第39期:198-202。
MOINARD, C.、MORISSE, J.P.和FAURE, J.M.(1998年)笼子面积、高度和栖木对于蛋鸡羽毛状况、骨折和死亡率的影响。《英国家禽科学》第39期:198-202。
NEWBERRY, R.C.、KEELING, L.J.、ESTEVEZ, I.和BILCíK, B.(2007年)年轻作为预测因素时成年蛋鸡的严重啄羽行为:重新审视觅食转向假说。《应用动物行为科学》第107期:262-274。
RODENBURG, T.B.、BUITENHUIS, A.J.、ASK, B.、UITDEHAAG, K.A.、KOENE, P.、VAN DER POEL, J.J.、VAN ARENDONK, J.A.M.和BOVENHUIS, H.(2004年)两个不同年龄段蛋鸡的啄羽和旷场反应之间的遗传和表型相关性。《行为遗传学》第34期:407-415。
RODENBURG, T.B.、DE HAAS, E.N.、NIELSEN, B.L.和BUITENHUIS, A.J.(2010年)选育啄羽表现强和弱的蛋鸡研究蛋鸡的恐惧和羽毛损伤。《应用动物行为科学》第128期:91-96。
SEFABAR(2003年)欧洲农场动物的可持续育种和繁殖。http://www.effab.info/sefabar.html。
VAN HIERDEN, Y.M.、KORTE, S.M.、RUESINK, E.W.、VAN REENEN, C.G.、ENGEL, B.、KORTEBOUWS, G.A.H.、KOOLHAAS, J.M.和BLOKHUIS, H.J.(2002年)啄羽表现强和弱的选育蛋鸡幼雏的肾上腺皮质反应性以及中枢血清素和多巴胺周转。《生理与行为》第75期:653-659。
VESTERGAARD, K.S.和LISBORG, L.(1993年)与家禽沙浴有关的啄羽发育模型。《行为》第126期:291-308。
WECHSLER, B.和HUBER-EICHER, B.(1998年)饲料和栖木高度对于蛋鸡啄羽和羽毛损伤的影响。《应用动物行为科学》第58期:131-141。