








Summary
数年来,通过酸化饮用水和饲料控制沙门氏菌。对于饮用水和粉末状饲料,选酸取决于其杀灭细菌的能力。目前,市场上还出现了经过酸涂布或浸渍处理的产品,旨在进一步降低胃肠道中酸的浓度。研究表明,在饲料中添加涂有乙酸的产品会增加沙门氏菌在鸡肠道中的定植,而添加涂有丙酸和丁酸的产品则会减少鸡肠道中沙门氏菌的定植。此外,研究表明,整个育成期直至屠宰日龄,对于肉鸡种鸡模型,添加涂有丁酸的产品可减少粪便脱落。
原因可能在于有机酸对于沙门氏菌致病性的影响,因为接触低浓度乙酸会增加沙门氏菌对于肠道上皮细胞的侵袭能力,而非生长抑制浓度的丙酸和丁酸会降低沙门氏菌的侵袭能力。
这是由于沙门氏菌致病岛I基因的致病基因表达发生了变化,而这种表达是沙门氏菌侵袭肠道上皮细胞时所必需的。目前,多个机构正在研究相应方法,以增加能够产生丁酸的肠道微生物。通过相应的营养策略和添加益生菌或益生元的方式提高家禽盲肠中能够产生丁酸盐的微生物浓度,将成为抗击沙门氏菌感染的有效方法。事实上,这些细菌会在沙门氏菌的主要定植和入侵部位(即盲肠)产生丁酸。此外,凭经验在家禽饲料中加入添加剂的方法已有多年的应用历史,而现在这种方法终于有了更为合理的目标,即增加鸡盲肠中能够产生丁酸盐的菌群。
引言
禽肉和禽蛋是重要的食品,对于欧盟农业产业,家禽相关产业是重要的经济组成部分。但该行业同样面临着压力和各种挑战。最重要的是,人们需要营养价值高,且没有微生物和化学危害的禽蛋和禽肉。但不幸的是,禽蛋和禽肉是人类食源性细菌感染的主要来源之一。在这方面,沙门氏菌(主要是肠炎沙门氏菌)和弯曲杆菌尤为危险,因为这些病原体可定植于鸡宿主体内,但不会导致感染的鸡产生明显病状。 在欧洲和美国,肠炎沙门氏菌是迄今为止最重要的禽蛋相关病原体,能够导致人类非伤寒沙门氏菌感染大流行。由于对蛋鸡群采取了控制措施,最近人类感染肠炎沙门氏菌的病例有所减少,但沙门氏菌仍然是世界上最重要的食源性细菌感染源之一。其他血清型的沙门氏菌发病率并未出现类似肠炎沙门氏菌的降低现象。 虽然有些血清型可追溯至猪肉,例如德尔比沙门氏菌和鼠伤寒沙门氏菌,但其他血清型则可追溯至禽肉,例如魏尔肖沙门氏菌、婴儿沙门氏菌、哈达尔沙门氏菌和鼠伤寒沙门氏菌。根据欧盟法律规定,从2010年12月12日起,如果禽肉中检测出沙门氏菌,将不能作为鲜禽肉上市销售,因此肉鸡行业需采取措施减少肉鸡及其所处环境中的沙门氏菌菌落。 包括屠宰前、屠宰中和屠宰后的相应措施。所有这些措施都同等重要,每项措施均或多或少对降低沙门氏菌发病率产生影响,单独采取某项措施不会获得相应的效果。屠宰中措施主要包括捕捉和运输过程中的卫生措施,而屠宰后措施则包括相应的卫生措施和肉类的净化处理。 然而,欧盟目前禁止使用所有类型的屠体消毒剂,因此无法进行净化处理。因此,在欧洲,活体阶段(屠宰前)的预防和监测/消杀工作非常重要。屠宰前的预防和控制措施始于专门的饲料厂和初级饲养者的祖代农场(MacLeod,2002年),直至肉鸡养殖者和鸡蛋生产商的商业农场,包括预防性卫生措施以及针对饲料、饮用水和禽类所处环境的物理和化学净化处理。由于抗生素不是控制沙门氏菌的首选方法,而且肉鸡的寿命较短,无法接种疫苗,因此将饲料添加剂作为细菌性肠道感染重要防治措施的做法得到了越来越广泛的应用。 但是,目前市场上有许多类似产品,且大多数产品仅凭经验生产,因此,家禽生产商很难客观地选出经过实践证实对于沙门氏菌有效的产品。使用有机酸抗击沙门氏菌
短链脂肪酸对于沙门氏菌体外存活力和致病性的影响对沙门氏菌,中链脂肪酸(C6至C12;己酸、辛酸、癸酸和十二酸)似乎比短链脂肪酸(甲酸、乙酸、丙酸和丁酸)更为有效。对于肠炎沙门氏菌菌株,只需25毫摩尔C6至C10酸即可实现抑菌效果,而这种菌株能够耐受100毫摩尔短链脂肪酸(Van Immerseel等,2003年,2004a)。肠炎沙门氏菌和鼠伤寒沙门氏菌在低浓度一癸酸甘油酯(5毫摩尔)环境中培养(通过饲料添加剂的合理设计预防沙门氏菌感染:第42(1)卷,2007年4月,第10页),细菌无法存活(Thormar等,2006年)。总的来说,这些数据表明,中链脂肪酸对于沙门氏菌的抗菌活性最强,但目前缺乏相应的大规模研究 。
沙门氏菌是一种机会性细胞内病原体,具有一系列复杂的致病基因。细菌能够凭借这些基因适应环境,并在宿主体内的不同微生境之间移动。沙门氏菌致病的第一步是穿透肠上皮。
位于致病岛(SPI-1)上的侵袭基因能够促进这种活性,但需要多个致病岛才能够实现完全的致病性。SPI-1拥有对于调节蛋白、针状复合体结构成分和其他效应蛋白具有编码作用的基因。通过诱导肌动蛋白基因重排(导致细菌吸收),细菌效应蛋白可促进沙门氏菌进入上皮细胞的细胞溶质。反过来,受环境调节的HilA蛋白能够激活SPI-1。如果在添加有不同浓度丁酸盐和丙酸盐的生长培养基中进行预培养,鼠伤寒沙门氏菌对于上皮细胞的侵袭会受到抑制(Van Immerseel等,2003年)。但是,如果细菌在添加有乙酸盐的培养基中进行预培养,仍然可观察到细菌的侵袭现象。
使用鸡的原发盲肠上皮细胞测试肠炎沙门氏菌,同样获得了类似结果。在SPI-1表达的变化中,能够解释有机酸对于上皮侵袭能力的影响。将鼠伤寒沙门氏菌暴露于pH值为6的乙酸盐、丁酸盐和丙酸盐环境,对于HilA和invF(SPI-1的主要活化剂)表达的测量结果显示,细菌暴露于乙酸盐环境会增加这些基因的表达,但丙酸盐和丁酸盐环境未观察到类似影响(Durant等,2000年)。
最近的研究表明,鼠伤寒沙门氏菌和沙门氏菌会降低hilA、invF和sipC的表达(Lawhon等,2005年)。肠炎沙门氏菌测试结果表明,低剂量丁酸会使SPI-1表达下调,但不会改变代谢基因的表达(Gantois等,2006年)。细菌细胞中的丁酸盐主要作用靶点尚不清楚,但丁酸盐可能会通过改变hilD转录调节的方式干扰SPI1的HilA依赖性调节。这些数据表明,短链脂肪酸对于沙门氏菌的侵袭表型具有调节作用。
短链脂肪酸对于体内沙门氏菌控制的影响
使用酸性化合物控制沙门氏菌的方法,最早出现于20世纪60年代末,主要用于骨粉饲料的净化。关于32种不同酸制剂的骨粉饲料净化效果的评估表明,低分子量挥发性脂肪酸最具应用前景(Khan和Katamay,1969年)。对于旨在控制沙门氏菌的无毒天然酸性化合物的开发工作,这些结果奠定了基础。35年过去了,很明显这些想法非常具有预见性。将这些酸类物质添加至饲料、饮用水和其他基质中,以防止沙门氏菌在动物组织中定植,同时防止其通过食物链传播。
家禽饲料是沙门氏菌侵入农场的主要源头。如果鸡摄入人为污染的饲料,鸡的肠道就会受到污染,沙门氏菌就会流入环境中。在饲料中添加酸类物质,最初的原因是酸类能够净化饲料本身,从而防止鸡吸收沙门氏菌。
WIba和Berchieri(1995年)在饲料中接种高剂量鼠伤寒沙门氏菌菌株,甲酸和丙酸的商用混合物能够在7天内将鼠伤寒沙门氏菌的存活率降低1000倍以上。肉鸡摄入混有肠炎沙门氏菌、鼠伤寒沙门氏菌或阿格纳沙门氏菌,而且经过酸处理的饲料,在第5天时,肉鸡盲肠中的沙门氏菌数量比对照组低7个对数级(102菌落形成单位/克对109菌落形成单位/克)。在饲料中人工接种低剂量凯道古沙门氏菌,甲酸和丙酸的混合物也很有效,储存数周后,沙门氏菌的减少现象最为明显。在一项大规模研究中,饲料中添加0.5%的甲酸后,沙门氏菌阳性的种鸡饲料样本比例从4.1%降至1.1%。(通过饲料添加剂的合理设计预防沙门氏菌感染:第42(1)卷,2007年4月,第11页)有机酸的抗菌活性取决于温度和湿度。由于家禽饲料的含水量通常较低,酸类物质的作用并不一定能达到最佳,产生保护作用的主要原因目前尚不清楚,无法确定是饲料中酸类物质本身发挥作用还是酸类物质在动物胃肠道中发挥作用。
20世纪80年代,人们发现嗉囊中的酸类物质浓度也在增加,这种抗菌作用有助于控制水平传播引起的感染。事实上,鸡摄入经过酸类物质处理的饲料后,饲料的温度和湿度均提升,短链脂肪酸的活性也同样增加。添加酸类物质的做法似乎最容易影响嗉囊和砂囊,而非肠道。Thompson和Hinton(1997年)的一项研究对此进行说明,在研究中蛋鸡摄入添加有甲酸和丙酸商用混合物的饲料。与对照组受试动物相比,这些动物的嗉囊、砂囊、空肠、盲肠和结肠pH值未发生变化,但嗉囊和砂囊中的甲酸和丙酸浓度显著增加。与此同时,嗉囊中的乳酸浓度明显下降,这表明乳酸杆菌受到了抑制或杀灭。
20世纪80年代后期,多项研究探讨了添加酸类物质的做法对于鸡组织沙门氏菌定植的影响。主要测试的是甲酸和丙酸的作用。在一项小规模的实地试验中,对于自然感染受试动物的沙门氏菌血清型变种,甲酸对于脱落和盲肠定植具有控制作用。事实上,50%的对照组受试动物泄殖腔拭子和盲肠内容物样本沙门氏菌呈阳性,但在摄入大量甲酸的受试动物体内却检测不到沙门氏菌(Hinton等,1985年)。
在一项为期3年的研究中,种鸡摄入经过甲酸处理的饲料,新孵化雏鸡感染沙门氏菌的累计数量有所减少(Humphrey和Lanning,1988年)。对于摄入酸化饲料的种鸡,便溺垫物(4.3%对1.4%)、孵化废弃物(15.3%对1.2%)和插纸样品(4.6%对1.4%)中脱落的沙门氏菌数量较少。自种鸡摄入酸化饲料起,下降现象尤为明显,证实了垂直传播的影响。
对于甲酸和丙酸作为饲料添加剂控制沙门氏菌的功效,Hinton和Linton(1988年)给出了最有力的证明。通过三项独立的实验,仅对自然感染的情况进行了监测。从孵化当天开始提供添加有甲酸和乙酸的饲料,可显著减少粪便和盲肠内容物阳性样本的数量。 对照组粪便样本的沙门氏菌阳性率分别为25%、27%和60%,而处理组分别为3%、0%和0%。如果在较晚日龄(16天或32天)喂食经过甲酸处理的饲料,对照组和处理组之间没有发现差异。这些结果说明,最重要的是防止沙门氏菌的初始定植。感染一旦形成,就很难通过经过酸类物质处理的饲料消除,至少同一生产轮次内是这样。
R最近,研究人员尝试通过微胶囊技术将有机酸进一步向胃肠道下游输送,以此防止上消化道吸收酸类物质,并确保酸类物质在胃肠道下游释放。Van Immerseel等(2004b)研究了含有甲酸、乙酸、丙酸和丁酸的微珠对于肠炎沙门氏菌盲肠、肝脏和脾脏定植的影响。利用5 x 103菌落形成单位的肠炎沙门氏菌感染受试动物(孵化后第5天),感染后3天采集相应的样本。饲料中添加乙酸时,盲肠定植率明显增加,而添加丁酸时,盲肠定植率下降(表1)。如果在饲料中添加甲酸或乙酸,内脏器官的定植率会增加,则这一结果与这些酸类物质能够增强沙门氏菌致病性的观点一致。
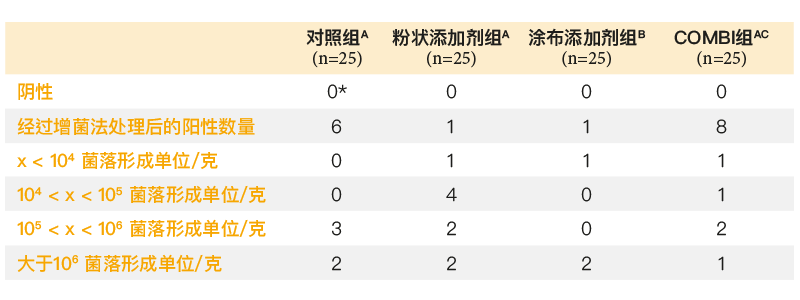
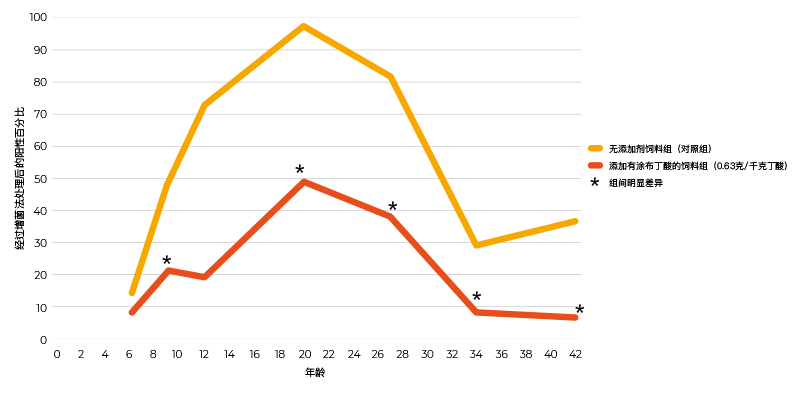
通过相同的感染方案对粉状和涂布丁酸添加剂(0.63克/千克丁酸)进行对比,涂布丁酸添加剂减少了盲肠的定植率,而粉状丁酸添加剂则没有发生变化(表2)。因感染和采样间相隔的时间间隔太短,粉状添加剂无法给出阳性反应。在一项采用种鸡模型的感染研究中,10只肉鸡在孵化后第5天感染105菌落形成单位的肠炎沙门氏菌,并与40只未接种肠炎沙门氏菌的肉鸡饲养在一起,饲料中添加0.63克/千克浓度涂布丁酸可显著减少肉鸡肠炎沙门氏菌的脱落,直至屠宰日龄(Van Immerseel等,2005年,(图1))。酸类物质对于微生物群落其他成员的影响尚未确定。
表1. 饲喂不同饲料(添加有甲酸、乙酸、丙酸和丁酸或不添加任何饲料添加剂)的鸡在孵化后第8天的盲肠定植情况(第5天和第6天接种103菌落形成单位的肠炎沙门氏菌76Sa88菌株)。
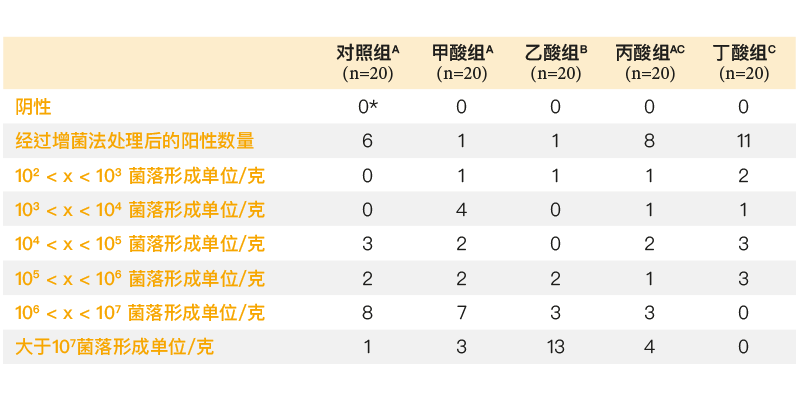
表2. 饲喂不同饲料(添加有粉状丁酸饲料添加剂、涂布丁酸饲料添加剂、粉状和涂布丁酸饲料添加剂半剂量混合物(COMBI)或无饲料添加剂(CTRL))的鸡在孵化后第8天的盲肠定植情况(第5天接种106菌落形成单位的肠炎沙门氏菌76Sa88菌株)。各组的活性产品丁酸浓度为0.63克/千克饲料。
通过提高盲肠丁酸盐浓度控制沙门氏菌:使用益生元和益生菌方法?
饲料和饮用水卫生以及在嗉囊中添加酸类物质,似乎可防止病原体在活体动物中定植,但酸类物质的种类和浓度非常重要。沙门氏菌在盲肠和内脏的定植并不总是受到这些处理方法的影响,尤其是在感染压力较高的情况下。因沙门氏菌主要在盲肠中定植,饲料或饮用水中的酸类物质在肠道的更深处不起作用。 由于盲肠是主要的发酵场所,短链脂肪酸的浓度已经高于其他肠段。各种动物物种实验已经表明 。
图1. 孵化后第5天10只雏鸡感染105菌落形成单位的肠炎沙门氏菌76Sa88菌株并与40只未接种沙门氏菌的肉鸡一起饲养后阳性泄殖腔拭子数量随着时间(日龄)变化的情况。
摄入双歧杆菌益生菌菌株或添加某些类型的低聚糖以刺激双歧杆菌在肠道中的增殖,如果双歧杆菌数量增加,沙门氏菌在肠道中的定植率则降低(Asahara等,2001年;Buddington等,2002年;Bovee-Oudenhoven等,2003年;Silva等,2004年;Thitaram等,2005年)。
肠道中乳酸菌数量的增加与丁酸浓度的增加相关,如果肠道中丁酸浓度增加,沙门氏菌的定植率则降低。双歧杆菌导致丁酸浓度增加,但双歧杆菌本身并不会产生丁酸。乳酸菌(例如乳酸杆菌和双歧杆菌)会刺激产丁酸细菌的增殖。这种机制称为交叉饲喂。
研究表明,厌氧链球菌accae和霍氏真杆菌(共培养)利用以淀粉为唯一碳源的青春双歧杆菌在体外产生的乳酸生产高浓度丁酸。另一种方法是直接刺激产丁酸细菌。在人类肠道样本中,丁酸的产生者是属于系统发育梭状芽孢杆菌群IV和XIVa的厌氧细菌,与罗氏菌属、真杆菌属、粪杆菌属和粪球菌属相关的物种也能够产生丁酸(Pryde等,2002年;Duncan等,2004年)。多种已发现的产丁酸菌群都是乙酸的净消费者。对于从鸡盲肠中分离出的16SrDNA序列进行随机克隆和测序,发现85%以上的克隆体属于真菌和梭状芽孢杆菌(Bjerrum等,2006年)。大约10%的克隆体与柔嫩梭菌(一种在人体肠道中产生丁酸的菌种)具有高度相似性。
作者的实验室从家禽盲肠中分离出能够产生丁酸的细菌(培养物中产生的丁酸超过10毫摩尔),16SrDNA序列分析表明,这些分离物与人类体内的产丁酸物种关系密切。这些分离物可消耗乙酸盐并产生丁酸盐,而且很有可能受到乳酸盐的交叉饲喂。这些数据有助于沙门氏菌控制技术的发展。事实上,通过相应的营养策略和添加益生菌或益生元的方式提高家禽盲肠中能够产生丁酸盐的微生物浓度,将成为抗击沙门氏菌感染的有效方法。
由于这些细菌会在沙门氏菌的主要定植和入侵部位产生丁酸,因此成功的几率会更大。此外,凭经验在家禽饲料中加入添加剂的方法已有多年的应用历史,而现在这种方法终于有了更为合理的目标,即增加鸡盲肠中能够产生丁酸盐的菌群,最终产生一种衡量添加剂作用的方法。因此,沙门氏菌的控制标准是:添加剂是否会增加鸡盲肠中能够产生丁酸的细菌的数量和分布?
目前正在开发一种检测方法,从DNA层面量化厌氧产丁酸菌群中参与丁酸合成的关键酶,以便在不进行大规模感染试验的情况下,通过简单可靠的检测方法评估添加剂和沙门氏菌控制的效果。这样有助于在未来实现饲料添加剂的合理设计。
Asahara, T.、Nomoto, K.、Shimizu, K.、Watanuki, M.和tanaka, R.(2001年)通过双歧杆菌和转半乳糖低聚醣的共生摄入提高小鼠对于鼠伤寒沙门氏菌肠道血清变种感染的抵抗力。《应用微生物学期刊》第91期,第985至996页。
Bjerrum, L.、Engberg, R.M.、Leser, T.D.、Jensen, B.B.、Finster, K.和Pedersen, K.(2006年)通过分子和培养技术揭示肉鸡回肠和盲肠微生物群落的构成。《家禽科学》第85期,第1151至1164页。
Bovee-Oudenhoven, I.M.、Ten Bruggencate, S.J.、Lettink-Wissink, M.L.、Van Der Meer, R.(2003年)通过饮食方式摄入的低聚果糖和乳糖可抑制沙门氏菌在大鼠肠道中的定植,但会刺激沙门氏菌的转移。《肠道》第52期,第1572至1578页。
Buddington, K.K.、Donahoo, J.B.和Buddington, R.K.(2002年)通过饮食方式摄入的低聚果糖和菊糖可保护小鼠免受肠道和全身性病原体和肿瘤诱导剂的侵害。《营养学期刊》第132期,第472至477页。
Duncan, S.H.、Holtrop, G.、Lobley, G.E.、Calder, A.G.、Stewart, C.S.和Flint, H.J.(2004年)人类粪便细菌产生丁酸盐过程中乙酸盐的作用。《英国营养学期刊》第91期,第915至923页。
Durant, J.A.、Corrier, D.E.和Ricke, S.C.(2000年)短链挥发性脂肪酸可调节鼠伤寒沙门氏菌hilA和invF基因的表达。《食品保护期刊》第63期,第573至578页。
Hinton, M.和Linton, A.H.(1988年)通过饲料的酸类物质处理控制肉鸡沙门氏菌感染。《兽医志期刊》第123期,第416至421页。
Hinton, M.、Linton, A.H.和Perry, F.G.(1985年)通过雏鸡饲料的酸类物质灭菌控制沙门氏菌。《兽医志期刊》第116期,第502页。
Humblot, C.、Bruneau, A.、Sutren, M.、Lhoste, E.F.、Dore, J.、Andrieux, C.和Rabot, S.(2005年)使用通用、乳酸杆菌和双歧杆菌16S rRNA基因引物进行PCR时相温度梯度凝胶电泳,结果显示抱子甘蓝、菊糖和发酵牛奶可改变人类微生物群相关大鼠的粪便微生物群。《英国营养学期刊》第93期,第677至684页。
Humphrey, T.J.和Lanning, D.G.(1988年)沙门氏菌的垂直传播与鸡饲料的甲酸处理。《流行病学与感染》第100期,第43至49页。
Iba, A.M.和Berchieri, A.(1995年)关于使用甲酸-丙酸混合物(Bio-Add™)控制肉鸡沙门氏菌感染(基于实验)的研究。《禽类病理学》第24期,第303至311页。
Khan, M.和Katamay, M.(1969年)脂肪酸对于肉骨粉中沙门氏菌的拮抗作用。《应用微生物学》第17期,第402至404页。
Lawhon, S.D.、Maurer, R.、Suyemoto, M和Altier, C.(2002年)肠道短链脂肪酸可通过BarA/SirA改变鼠伤寒沙门氏菌侵袭基因的表达和致病性。《分子微生物学》第46期,第1451至1464页。
Pryde, S.E.、Duncan, S.H.、Hold, G.L.、Stewart, C.S.和Flint, H.J.(2002年)人类结肠中丁酸盐形成的微生物学机制。《FEMS微生物学快报》第217期,第133至139页。
Thitaram, S.N.、Chung, C.H.、Day, D.F.、Hinton, A.Jr.、Bailey, J.S.、Siragusa, G.R.(2005年)异麦芽低聚糖可增加幼年肉鸡盲肠中双歧杆菌的数量。《家禽科学》第84期,第998至1003页。
Thompson, J.L.和Hinton, M.(1997年)母鸡饲料中的甲酸和丙酸对于嗉囊中沙门氏菌的抗菌活性。《英国家禽科学》第38期,第59至65页。
Thormar, H.、Hilmarsson, H.和Bersson, G.(2006年)对于食源性细菌空肠弯曲杆菌、沙门氏菌属和大肠杆菌具有抗菌活性的癸酸 1-单酸甘油酯(一癸酸甘油酯)稳定浓缩乳剂。《应用与环境微生物学》第72期,第522至526页。
Gantois, I.、Ducatelle, R.、Pasmans, F.、Haesebrouck, F.、Hautefort, I.、Thompson, A.、Hinton, J.C.和Van Immerseel, F.(2006年)丁酸盐对于沙门氏菌致病岛I基因的表达具有特异性下调作用。《应用与环境微生物学》第72期,第946至949页。
MacLeod, H.(2002年).饲料净化的饲养者实践经验。《罗曼家禽育种公司国际情报资料》第26期,第7至8页。
Silva, A.M.、Barbosa, F.H.、Duarte, R.、Vieira, L.Q.、Arantes, R.M.和Nicoli, J.R.(2004年)长双岐杆菌的摄入对于小鼠沙门氏菌病(基于实验)的影响。《应用微生物学期刊》第97期,第29至37页。
Van Immerseel, F.、De Buck, J.、Meulemans, G.、Pasmans, F.、Velge, P.、Bottreau, E.、Haesebrouck, F.和Ducatelle, R.(2003年) 肠炎沙门氏菌对于禽类肠道上皮细胞的体外侵袭受到短链脂肪酸的影响。《食品微生物学国际期刊》第85期,第237至248页。
Van Immerseel, F.、De Buck, J.、Boyen, F.、Bohez, L.、Pasmans, F.、Volf, J.、Sevcik, M.、Rychlik, I.、Haesebrouck, F.和Ducatelle, R.(2004a)鸡感染肠炎沙门氏菌后短期内中链脂肪酸可通过抑制hilA的方式减少肠炎沙门氏菌的定植和侵袭。《应用与环境微生物学》第70期,第3582至3587页。
Van Immerseel, F.、Fievez, V.、De Buck, J.、Pasmans, F.、Martel, A.、Haesebrouck, F.和Ducatelle, R.(2004b)幼鸡感染肠炎沙门氏菌早期饲料中的微胶囊短链脂肪酸可改变肠炎沙门氏菌的定植和侵袭情况。《家禽科学》第83期,第69至74页。
Van Immerseel, F.、Boyen, F.、Gantois, I.、Timbermont, L.、Bohez, L.、Pasmans, F.、Haesebrouck, F.和Ducatelle, R.(2005年)在饲料中添加涂布丁酸添加剂可减少家禽体内沙门氏菌的定植和脱落。《家禽科学》第84期,第1851至1856页。