










Summary
Drinking water for animals is a feedstuff and the most important nutrient. A shortage of water in the
organism quickly leads to a decrease in feed consumption, a reduction in growth, disturbances in
metabolism and finally to death. Biological and chemical ingredients can affect the health of the
animals and the consumer.
The importance of drinking water for the performance and health of the animals is often underestimated. The article addresses the issue of regulating the water balance of the animals, the water
requirements, drinking system technology, water quality and the maintenance and monitoring thereof,
with particular emphasis being placed of broiler chickens. It further deals with the possibility of administering various substances and preparations via drinking water. Knowledge gained at a workshop
on the subject „Water supply in poultry farming – a key factor for health and performance“ is included
in the article. The workshop with international guests was organised by Lohmann Animal Health and
took place in October 2009.
Introduction
Crossbreeding is a standard practice in poultry breeding programmes as a way of exploiting heterosis. However, there is no consensus on the most effective way to maximize the genetic response in crossbred commercial animals (Besbes and Ducrocq, 2003). Furthermore, the goal of breeding is not to maximise heterosis, but to maximise overall profitability in the commercial cross, the parents and the pure lines (Flock et al., 1991). Environmental differences exist between breeding farms (e.g. with single cages) and commercial farms. Thus, both environments should be taken into account in the breeding program, to minimise possible genotype-environment-interactions that can reduce the response to selection on the commercial level. Considering cross-bred and pure-bred performance as two separate, but correlated traits offers an elegant way to take these environmental effects into account. The cross-bred performance is captured in the environment in which the breeding goal is defined (Wei and van der Werf, 1995). In order to optimise a breeding program, ranking based on pure-bred performance has to be compared with ranking based on cross-bred performance on commercial farms. If there are significant differences, cross-bred information has to be included in the selection process. The aim of this study was to estimate genetic correlations between pure-bred and cross-bred performance in commercial white-egg and brown-egg lines as essential information to optimise the breeding programs.Material and methods
The pure-line data used for the present analysis represent three generations of a White Leghorn (WL) and a Rhode Island Red (RIR) male line, recorded in breeding farms with single cages and high standards of bio-security, feed quality and environment control. The single-cross daughters of the same sires were housed in group cages (4 hens per cage) in four commercial farms each generation, two farms for each cross. The cross-bred hens had only sire pedigree. On average, about 70 sires per line and generation were used to produce 50 WL and 69 RIR pure-bred daughters and 30 cross-bred daughters per line and generation.
The traits used for this study were cumulative early egg production between 20 and 27 weeks of age (P1), peak production between 28 and 47 weeks of age (P2) and late egg production between 48 and 68 weeks of age (P3). Average egg weight was measured at 29, 34 and 45 weeks of age (EW).
Furthermore, the average shell breaking strength (SS) was measured at 40, 43 and 46 weeks of age. Shell colour (SC) was measured at 30 and 45 weeks of age and averaged. Mortality was taken into account for egg production in group cages, dividing production per cage by the number of hens alive in each period. In the cross line egg quality traits (EW, SS and SC) were recorded at one day’s production, and cage means were calculated from individual measurements. Only cages with 4 hens at the beginning of production were included in this study to avoid heterogeneity of residual variance of multiple bird cage data due to different group size.
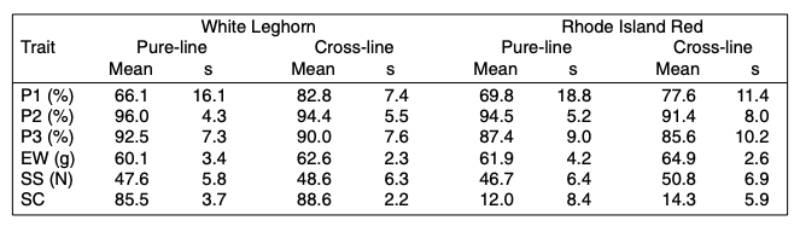
A total of 10,002 and 13,367 pure-bred records and 1,426 and 1,394 cage averages for the WL and RIR cross-breds were included in the statistical analysis. Table 1 shows an overview of the general statistics for the different traits.
The program VCE 4 (Neumaier and Groeneveld, 1998) based on the REML method and a multiple trait animal model was used to estimate (co)variance components, with the model:
yijk = µ + GHHTi + aj + eijk
where yijk is the vector of observations for the different traits; µ is the population mean; GHHTi is the combined fixed effect of generation, house, hatch and tier; aj is the random additive genetic effect of
Table 1: Means and standard deviations for the traits studied
animal j; and eijk is the random residual effect. In the case of the cross-line data for laying rate, only a cage average could be measured and only the sire was known. Therefore, in the model the cage mean was used as vector of observations, and only one hen out of the half-sib family cage was used for the animal effect. Theoretically, the residual variance estimated from pooled observations should be about n times the estimate based on individual observations (Biscarini et al., 2008). According to this author, the heritability for pooled data should be recalculated as: h2 = σa 2 / (σa 2 + n σe 2); where ‘n’ is the number of hens/cage (4 in this case). This is in line with Simianer and Gjerde (1991) who explained that estimation based on group means contains less information on the group variation, which essentially contributes to the estimation of residual variance components.
Results and discussion
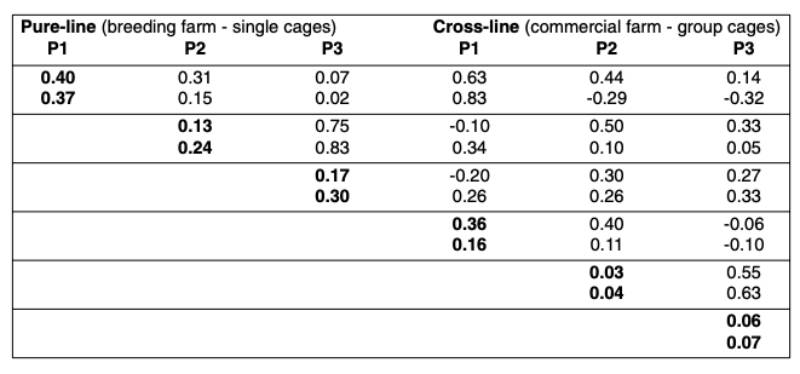
Table 1 shows the means for all traits included in the analysis. Most people would expect higher performance in single crosses than in the pure-breds due to heterosis, but as shown in the table this is not always the case. It should be borne in mind that the pure-lines were kept under optimal biosecurity and management conditions, whereas the cross-breds were kept under less ideal commercial environment in multiple-bird cages.Table 2 shows the heritabilities and genetic correlations. Early egg production (P1) had the highest heritability, which is explained by variation in age at sexual maturity (Preisinger and Savas, 1997).
The heritability was lowest during peak production and increased again towards the end of lay, confirming estimates from the literature (e.g. Anang et al., 2000). No or a slightly negative correlation was found between the production at sexual maturity and at the end of lay. As expected, the genetic correlation was high between peak production and persistency.
The heritabilities were consistently higher based on full pedigree pure-line records from single cages than based on the cross-line data from commercial multiple bird cages. The genetic correlations between pure-bred and cross-bred performances were moderate to high at sexual maturity (rg = 0.63 and 0.83), whereas the correlations decreased to moderate to low levels in the other two stages of production (rg between 0.1 and 0.5). The heritabilities obtained in this study were similar to those published by Nurgiartiningsih et al. (2004).
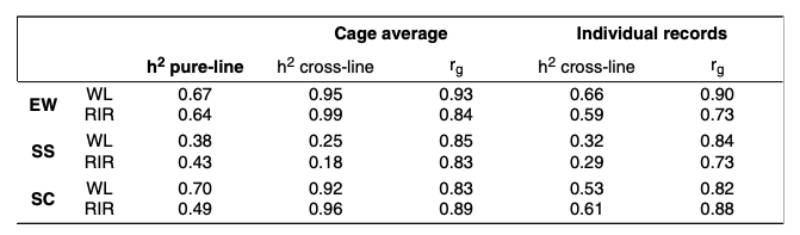
Estimates of genetic parameters for egg quality traits are summarized in table 3. For the cross-breds, the individual egg quality measurements were used. Additionally, cage averages were assigned to one of the hens in the cage and used for the analysis to underline the problems of using these traits as a cage average. The heritability estimates based on cage averages are obviously too high and reflect the reduced residual variance when using cage averages. Using the single egg records, the heritability estimates are slightly lower than in the pure-breds, which would be expected under « suboptimal » conditions. It should also be noted that only one egg per hen was evaluated for group cages, whereas several eggs per hen were evaluated for the pure-lines in single cages.
Table 2: Genetic Parameters for egg production traits in pure-line and cross-line (cross-line
data as cage average, WL in the first row and RIR in the second)
Estimates of genetic parameters for egg quality traits are summarized in table 3. For the cross-breds, the individual egg quality measurements was used. Additionally, cage averages were assigned to one of the hens in the cage and used for the analysis to underline the problem of using these traits as cage averages. The heritability estimates based on cage averages are obviously too high and reflect the reduced residual variance when using cage averages. Using the single egg records, the heritability estimates are slightly lower than in the pure-breds, which would be expected under “suboptimal” conditions. It should also be noted that only one egg per hen was evaluated for group cages, whereas several eggs per hen were evaluated for the pure-lines in single cages.
Table 3: Genetic Parameters for egg quality traits in pure-lines and cross-lines, with crossbred data as cage average and as individual records
Wei et al. (1995) concluded that combined cross-bred and pure-bred selection should be better than selection based only on pure-line performance when improvement of cross-bred performance is the goal. This strategy is generally more efficient when genetic correlations are below 0.8, which is the case for laying rate. However, it is essential to find suitable farms with reliable data recording and flexible housing schedules to match well-designed tests to augment the basis for selection focused on the needs of the egg industry. If the residual variance is high due to uncontrolled factors which are not representative for other farms, the heritability will be low and the gain from using this information questionable.
Conclusions
The results of this study indicate that the genetic correlation between pure-line and cross-line egg production was moderate, whereas the genetic correlations for all egg quality traits were high. This confirms the working hypothesis that egg production data of cross-line relatives collected under commercial conditions can contribute significantly to genetic improvement, while selection for egg quality traits can be limited to pure-line records. The loss of statistical information on individual variation due to working with cage mean is inevitable, but group cage data of cross-lines can help to reduce the effect of genotype-environment interaction while taking heterosis effects into account. The parameter estimates found in this study confirm that combined pure-line and cross-line selection is an efficient approach to improve total genetic merit under field conditions.Zusammenfassung
Genetische Parameter von Reinzucht- und Kreuzungsleistungen bei Legehennen Mit einem Mehrmerkmals – Tiermodel wurden genetische Parameter für Reinzucht- und Kreuzungsleistungen geschätzt. Die Daten wurden über drei Generationen von zwei kommerziellen Reinzuchtlinien in Einzelkäfigen und Kreuzungsnachkommen derselben Väter in Gruppenkäfigen erfasst. Die Heritabilitäten für Legeleistung liegen im Bereich von h2=0,03 bis 0,40). Die genetischen Korrelationen zwischen Reinzucht- und Kreuzungsleistung nehmen im Verlauf der Produktion von anfangs rg= +0,63 bis +0,83 auf rg= +0,10 bis +0,50 ab und unterstreichen damit die Notwendigkeit, unter praxisnahen Bedingungen auf Persistenz der Legeleistung zu testen. Die untersuchten Eiqualitätsmerkmale haben in beiden Umwelten eine mittlere bis hohe Heritabilität und durchweg hohe genetische Korrelationen zwischen Reinzucht- und Kreuzungsleistung.ANANG, A., N. MIELENZ, L. SCHÜLER (2000): Genetic and phenotypic parameters for monthly egg production in White Leghorn hens. J. Anim. Breed. Genet. 117, 407-415.
BESBES B., V. DUCROCQ (2003): Use of mixed model methodology in breeding strategies for layers. Poultry Genetics, Breeding and Biotechnology. Ed. by W.M. Muir and S.E. Aggrey. CABI Publishing, Wallingford, UK.
BISCARINI, F., H. BOVENHUIS, J.A.M. VAN ARENDONK (2008): Estimation of variance components and prediction of breeding values using pooled data. J. Anim. Sci. 86, 2845-2852.
FLOCK, D.K., H. AMELI, P. GLODEK (1991): Inbreeding and heterosis effects on quantitative traits in a white leghorn population under long-term reciprocal recurrent selection. Brit. Poult. Sci. 32, 451-462.
NEUMAIER, A., E. GROENEVELD (1998): Restricted maximum likelihood estimation of covariance in sparse linear models. Genet. Sel. Evol. 30, 3-26.
NURGIARTININGSIH, V.M.A., N. MIELENZ, R. PREISINGER, M. SCHMUTZ and L. SCHÜLER (2004): Estimation of genetic parameters based on individual and group mean records in laying hens. British Poultry Science 45, 1-7. PREISINGER, R., T. SAVAS, (1997): Vergleich zweier Methoden zur Schätzung der Varianzkomponenten für Leistungsmerkmale bei Legehennen. Züchtungskunde 69, 142-152.
SIMIANER H., D. GJERDE (1991): Estimating variance components from fullsib group means. J. Anim. Breed. Genet. 108, 270-279.
WEI, M., J.H.J. VAN DER WERF (1995): Genetic correlation and heritabilities for pure-bred and cross-bred performance in poultry egg production traits. J. Anim. Sci. 73, 2220-2226.