
Summary
Crossbreeding in dairy cattle is of growing interest to dairy cattle breeders and milk producers since
in the dominant dairy breed, i.e. Holstein, negative phenotypic trends have been observed for functional
traits like health, fertility and longevity. Although Holsteins are the most profitable dairy cows on well
managed farms, this may not apply to other farms. Crossbreeding to exploit heterosis effects for lowly
heritable functional traits is a tempting alternative. Under specific production systems, e.g. in New
Zealand, where high stocking rate and production of solids per hectare is the primary goal, fertility
and survival are even more in the focus and hence a significant share of the New Zealand dairy cows
population are indeed crossbreds.
A review of literature indicates that heterosis effects of 4 to 5 % for production traits and substantially more heterosis for fertility and survival can be expected. Recent experimental data from Germany indicate that the advantage of crosses is negligible at a very high management level. For some traits, e.g. feed efficiency, even adverse effects can be observed. At a lower level of management, crosses exhibit their superiority with respect to production, fertility, and survival. Unresolved questions remain:
a) the optimal design for a long-term crossbreeding system, assuming that farmers prefer present practices to produce their own replacements and
b) the apparent need to progeny-test individual bulls of ‘sire’ breeds for combining ability with Holstein cows.
A review of literature indicates that heterosis effects of 4 to 5 % for production traits and substantially more heterosis for fertility and survival can be expected. Recent experimental data from Germany indicate that the advantage of crosses is negligible at a very high management level. For some traits, e.g. feed efficiency, even adverse effects can be observed. At a lower level of management, crosses exhibit their superiority with respect to production, fertility, and survival. Unresolved questions remain:
a) the optimal design for a long-term crossbreeding system, assuming that farmers prefer present practices to produce their own replacements and
b) the apparent need to progeny-test individual bulls of ‘sire’ breeds for combining ability with Holstein cows.
Introduction
Crossbreeding has been the breeding method of choice for several species in plants and livestock. In plants, the main reason for crossbreeding is to exploit heterosis effects which can be maximized via the use of highly inbred parental strains. In livestock, notably in poultry and swine, positional effects of pure-bred parents of crosses are well known and hence crossbreeding systems are set up accordingly. Compared to plant breeding, heterosis effects are not expected to be as high but are still significant and thus give an additional benefit. Breeding systems for swine and poultry thus almost exclusively use crossbreeding systems.In dairy cattle, however, pure breeding has been the dominant breeding method for many years with only few exceptions. The reasons for this, comparing dairy cattle to swine or poultry, are mainly due to biological facts: Dairy cows have a much lower reproduction rate (≤ 1 offspring per year), a long generation interval and the individual animal has a relatively high value. These facts have contributed to a situation in which little difference is observed between commercial milk producers and pedigree breeders. In most European countries, large fractions of the populations are also registered as breeding stock, whereas in the United States commercial herds without any registered cattle exist besides herds with varying fractions of registered cattle.
Throughout the world, the main activities of breeding organizations still rely heavily on selecting animals from the entire population of a country or region. Some exceptions from this rule also exist. These pertain to organizations maintaining their own nuclei. However, these organizations have not yet captured a significant share of the world markets for bull semen, frozen embryos or live cattle. During the Eighties, dairy cattle breeders worldwide explored possibilities arising from the use of new biotechnologies to overcome the ineffectiveness of conventional breeding systems. One of these biotechnologies was embryo transfer including deep-freezing of embryos. However, initial progress levelled off with still unsatisfactory success rates. Around seven transferable embryos per flush after super-ovulation for cows reacting properly to a super-ovulation (equal to four pregnancies) is a level which has given embryo transfer only limited significance for increasing the reproductive rate of highly selected females. For the broad population, however, the significance of embryo transfer is close to nil.
Sexing of bull semen with sufficient accuracy has finally become available for dairy cattle breeders and for commercial milk producers worldwide. However, due to the high cost of semen sexing and low pregnancy rates this method so far is being used only in niche parts of the entire bull semen market.
Conventional and improved systems of purebreeding
Most conventional systems rely on selecting potential dams of bulls in the field, i.e. from herds owned by individual breeders. These bull dams are then mated to highly selected proven bulls. Their male offspring is reared for about one year until reaching sexual maturity, semen is collected from these young bulls and subsequently used in appropriate volume (400 to 1000 doses) for a progeny test under field conditions. The female offspring from these matings is reared to obtain cows with first calving and first lactation milk yield. Based on the results obtained from the milk recording systems in the field, finally, around four to five years after birth of the young bull, breeding values for dairy production can be estimated for these bulls. Based on the results of the genetic evaluation, selection can take place. Most bulls are slaughtered (around 90 %), and the remaining ones can be marketed by the breeding organization as proven bulls. Within the marketable proven bulls, the few very best are again selected for the elite matings with potential bull dams.Modifications of this basic scheme have included the use of embryo transfer for bull dams in order to increase the probability that male calves are born. Some attempts for closed nuclei have also been made. The advantages of nucleus breeding schemes in dairy cattle obviously are the direct control over all breeding activities by the breeding organization, the facilitation of accurate recording of traits other than production traits and logistic advantages when implementing biotechnologies. However, systems trying to keep nucleus lines fully closed without progeny testing young bulls in the field have failed. Keeping nucleus lines closed under a dairy cattle situation has the distinct disadvantage that the size of such operations is too small and hence possibilities to find genetically superior animals in the broad population still exist and should be exploited. Relying only on sib-testing within a small nucleus population will lead to breeding values of low reliability and consequently less variation compared to bulls progeny-tested in the field. Therefore, nucleus schemes still exist but are usually kept open and also use a progeny test in the broad population. In the future, these conditions may change if and when genomic selection based on large Single Nucleotide Polymorphisms chips (SNP) becomes the method of choice.
Growing interest in crossbreeding
In dairy cattle, the Holstein breed, formally called Holstein Friesian, has become the dominant breed throughout the world. Today, Holstein cows have the genetic potential to produce >10,000 kg milk per cow and lactation with a protein content of around 3.3 to 3.5 %. Given adequate nutrition, housing and health care, this level of production is combined with acceptable performance in functional traits such as fertility, health, functional conformation, and longevity. Genetic antagonisms between dairy production traits and functional traits exist but are usually low, within a range between zero and -0.3. Given this, genetic trends of most functional trends are slightly negative and nowadays breeding organizations focus on this problem by giving more and more weight to functional traits within the aggregate genotype. Most total merit indices used by the major breeding organizations today use weights of around 50 % for production traits and 50 % for functional traits.On the phenotypic level, however, many dairy operations fail to manage their high producing Holstein cows adequately and thus phenotypic trends for functional traits in general appear more negative than genetic ones. Examples are stillbirths and mastitis resistance. Well managed farms achieve results with < 3 to 4 % stillbirths and < 20 % mastitis cases per cow and lactation. For less well managed farms, stillbirth rates of > 10 % and mastitis cases > 50 % are common. In view of this situation, dairy farmers are increasingly looking for new ways to overcome these difficulties which directly affect their family income. Since health and longevity are traits with known low heritability, heterosis effects can be expected from specific crosses, supporting the growing interest in crossbreeding dairy cattle.
Crossbreeding in practice
For production conditions with high costs for labour and building, optimal economic returns are achieved when maximizing production per cow. This is the case for most parts of Europe as well as for North America. In New Zealand, on the other hand, barns to house the cows are not needed or can be very simple, because the climate permits year-round grazing. The optimal production system in this case includes strict seasonal calving and a dry period during the southern winter. Under such conditions, economic returns are maximized with increased stocking rate per area of grassland and therefore the focus in New Zealand is on maximizing milk solids per hectare. Herd production records averaged 3,736 kg milk, 186 kg fat and 139 kg protein per cow in 2005/06 (LIC, 2006) and thus are very low compared to North American or European statistics. On the other hand, New Zealand achieves a stocking rate of 2.8 cows per hectare and hence a production of 520 kg fat per effective hectare. Although statistics based on this parameter are not available for other countries, it can be assumed that the latter figure presents a world record. The high stocking rate and strict seasonal calving emphasize that a focus on survival and fertility is needed for New Zealand’s dairy producers and explains why crossbreeding and the exploitation of heterosis effects are common practice. Among the New Zealand cows under milk recording, 47 % belong to the Friesian (Holstein) breed, 17 % are Jersey, 1 % are Ayrshires and 35 % are crosses between Holstein and Jersey (LIC, 2006).Although the Holstein breed under North American and European conditions nowadays is capable of herd averages > 10,000 kg milk per cow, under New Zealand conditions the advantage in dairy production over other breeds or crosses on a per cow basis is only slight if not zero. However, for the lowly heritable traits survival and fertility, crosses show marked heterotic effects and hence are a natural choice under such conditions. On a per hectare basis, crosses are superior over pure breeds.
To a large extent, crossbreeding has also been done in beef cattle. The explanation for this is similar to the case of New Zealand for dairy cattle: A beef cow is expected to produce one weaned calf annually, and this is almost all a commercial producer expects from her. Fertility and survival is the focus for the commercial beef cattle ranger while pedigree breeders also try to improve fattening abilities and carcase traits. For many commercial producers, it is therefore quite logical to work with a crossbred cow. In many European countries, commercial beef cattle holders often start their business with crossbreds between dairy breeds and beef breeds. This system ensures that the crossbred dam not only exhibits heterosis effects with respect to fertility and survival, but also produces more milk for her calf than any beef breed.
Results from the literature
Reviewing the literature, Touchberry (1992) points out that a number of crossbreeding experiments in dairy cattle actually were carried out in the first half of the 20th century. A long term experiment involving the Holstein and Guernsey breeds was carried out in the 50’s in Illinois. Heterosis amounted to between 5 and 8 % for production traits and 12.8 % and 9.4 % for number of inseminations per pregnancy and days open, respectively. This underlines the potential for exploiting heterosis effects for fertility. In another publication, McAllister et al. (1994) presented results from a crossbreeding experiment involving Holstein and Ayrshire. For ‘annualized discounted net return’, the ‘bottom line’ trait from an economic perspective, 20 % heterosis was achieved. Some of the crossbred combinations were superior over purebreds in production, but the advantage of crossbreds was mainly due to superior fertility and survival.Given the above as outlined in the previous section, Lopez-Villalobos et al. (2000abc) found positive heterotic effects for virtually all traits of importance to New Zealand’s dairy producers and estimated a 25 % advantage for a rotational cross between Jersey and Holstein over the purebreds for the trait net income per hectare.
Van Raden and Sanders (2001) used the large US data base for the genetic evaluation of all dairy breeds and identified the various crosses present in the data. It should be noted that the vast majority of their data represented purebreds. Applying statistical models they estimated heterosis effects of 3 to 4 % for milk production traits, but were unable to detect any heterosis for somatic cell count, an important trait to dairy producers. Furthermore, heterosis effects for length of productive life were only at a level of 1.2 %.
Recently, Heins et al. (2006abc) reported on the analysis of crossbreeding data from seven large dairies in California. Apart from the Holstein breed which was used by all dairies, the breeds Normande, Montbeliarde, and Scandinavian Red (SR) were used for crossbreeding. The latter breed actually consisted of two breeds, the Norwegian Red and the Swedish Red Breed (SRB). Production of the crossbreds was lower than for Holsteins, but not statistically different for total fat plus protein yield of SR crosses. Significant advantages for crossbreds and especially for SR crosses were observed for calving ease, stillbirths, fertility and survival. The results from these California data have led to a very significant demand for semen of SRB bulls in the USA and in Europe to be used for crossbreeding (Svensk Avel, pers. comm., 2007).
Results from own data in Germany
Brown Swiss x Holstein in a high yielding herd
The results on crossbreeding reported in the literature are typically based on field data without underlying experimental design. Obviously, a planned experiment on crossbreeding in dairy cattle is costly, of limited size, or both. We attempted to carry out a designed experiment involving Holstein purebreds (HOL) in comparison with Brown Swiss x Holstein crosses (BS x HOL) to make up for limitations in size with accurate planning and repeated measurement of a large number of traits per animal. The experiment, carried out in cooperation between the State Research Agency for Agriculture of the state of Saxony Anhalt and the University of Halle, is still in progress and involves the high yielding herd of the State Research Agency in Iden. In 2003, when the first cows of the experiment calved, this herd (376 cows) had an average production of 11,186 kg milk, 422 kg fat, and 380 kg protein per cow.Two groups were formed, a Holstein purebred group and a crossbred group. For both groups, 10 sires each were selected involving sires of high genetic merit for both breeds and available in Germany in 2002. A total of 355 inseminations were carried out with 10 to 15 inseminations per sire. Sires were mated to cows well distributed over parities and genetic merit as estimated by cows’ breeding values. 134 calves were born in both groups from 124 (BS x HOL) and 131 (HOL) pregnancies, respectively. Deducting losses during birth and during rearing, and deducting male calves which were sold after birth, 55 BS x HOL and 51 HOL heifers calved and started lactating. Early losses during the first lactation left 50 BS x HOL and 45 HOL first calf cows with complete lactation records that could be evaluated using a ‘test day model’, in our case
From the Iden experiment: A Holstein cow flanked by three Brown
Swiss x Holstein crosses.

While all cows have completed their first lactation, most of them are in their second lactation and a few have begun their third lactation, we can summarize the first preliminary findings as follows:
- BS x HOL calves had higher birth weights compared to HOL. Also, the sexual dimorphism was more pronounced. No increases in calving problems were observed since calves with extreme birth weights, mostly BS x HOL in the range of 50 to 55 kg, were almost exclusively born from older dams.
- Weights and backfat thickness during time of rearing throughout the rearing period were equal for both groups.
- Additive effects due to the crossbreeding partner BS as sire breed were observed for measurements taken from the claw (more ‘compact’ and upright for BS x HOL) and taken from the udder (longer and thicker teats for BS x HOL).
- No differences were observed for calving ease and stillbirth for births of the cows of both groups in the first parity.
- Milking speed for BS x HOL cows was significantly slower than for HOL.
- Metabolic parameters measured from milk and urine were within a ‘healthy’ range for both groups although significant differences existed, e.g. for urea content of milk and urine (increased in BS x HOL).
- BS x HOL cows tended to show an improved fertility, but the difference was non-significant in the first lactation.
- Daily feed intake from day 7 to day 56 after calving was higher for BS x HOL (17.00 kg) than for HOL, which consumed 16.14 kg drymatter per day.
Table 1: Results (LSMEANS) for production traits (production per cow and day) from the
Iden experiment for Holstein purebreds (HOL) and Brown Swiss x Holstein (BS x
HOL) crosses (asterisks denote significant differences)
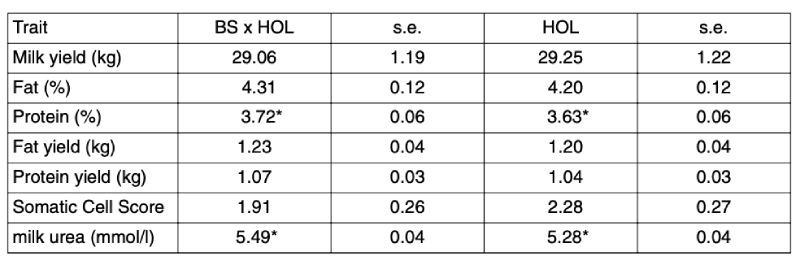
Summarizing the results, at present, no clear advantage of crossbreds can be seen from this experiment. Quite contrary, the inferiority of BS x HOL for the important traits milking speed and especially feed consumption would lead to the preliminary conclusion to rather stick with purebreds than BS crossbreds. However, the experiment is still in progress and differences in survival after several lactations as well as fertility parameters from several lactations have to be analyzed before drawing final conclusions. Furthermore, the preliminary conclusion may only hold for a high yielding dairy herd with close to perfect management as given at Iden.
Swedish Red Breed x Holstein and Brown Swiss x Holstein on an organic farm
Since 2002, the owner of a large organic farm in the state of Brandenburg has been crossbreeding with bulls of the Swedish Red Breed and also with Brown Swiss bulls on the basis of his original Holstein herd. The herd average under the organic conditions is around 7500 kg of milk per cow and lactation.

As for other reports in the literature, the data obtained from the farm are not from a planned experiment and represent the specific situation and genetic strategy on the farm. So far, comparisons can be made for Holstein purebreds, SRB x HOL crosses and BS x HOL crosses. No purebreds of the BS and SRB breeds are available on the farm and thus heterosis effects can not be estimated. Data from calving years 2003 to July 2007 were used for the analysis. During this time, substantial data were available for Holstein purebreds and the two crosses SRB x HOL and BS x HOL. Other crosses, e.g. HOL x (SRB x HOL) and BS x (SRB x HOL) were also present in the data but still of a limited number so that the results will not be presented here. The breeding strategy of the farm now is a true threeway rotational cross between the three breeds mentioned. Therefore, future data, once the lactations have been completed, will be available and subject to further analysis.
For the two crossbred groups, four sires of the breeds SRB and BS were used, respectively. This fact underlines that the results should be interpreted with great care since four sires per breed hardly can be viewed as a representative sample of the breed.
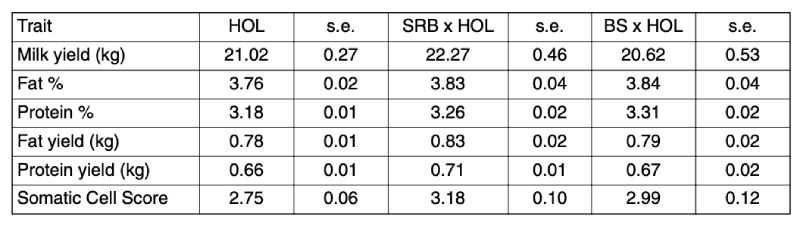
The number of cows by breed group and the number of test day records available for analysis under a test day model including regression factors for the curve of the lactation, age at calving, breed group, and cow within breed group is given in Table 2. The results for production traits are presented in Table 3.
SRB x HOL crosses show distinct and significant advantages over the two other breeds for all traits except protein percentage and somatic cell score. For protein percentage, the additive genetic effect of the Brown Swiss breed was expected to yield superior results in the crosses. The somatic cell scores are within a ‘healthy’ range for all three groups and thus observed differences may not be relevant in practice, even if statistically significant.
Table 2: Number of animals and test day records by breed group (Brandenburg data)
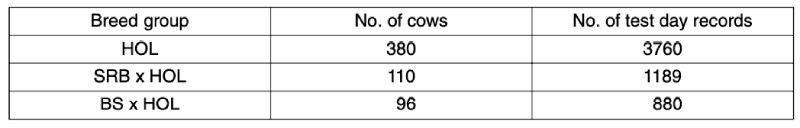
Table 3: Production characteristics (LSMEANS, production per cow and day) for the three
breed groups in the Brandenburg data
Since the number of sires for the crossbred groups was limited, a further test day model was fitted including sires as fixed effects. From this analysis, sires were ranked and the rankings were compared to their estimated breeding values in the respective breed of origin. Not too surprisingly, ranks of sires differed between performance in purebreds and crossbred animals. For BS sires, one striking example was a highly significant difference of 5 kg milk per day in the progeny of two bulls that had equal breeding values for milk production in their respective purebred population. This finding gives a hint
that a specific combining ability not only may exist for a specific cross but also for individual bulls. The three breed groups were also compared with respect to survival and fertility. For survival, losses within the first 365 days after the first calving were compared. HOL had losses of 16.8 %, SRB x HOL showed only 8.2 % and BS x HOL equalled 18.1 %. For fertility, the traits days to first service (DFS) and number of inseminations per pregnancy (NIPP) were considered. No significant differences were found for DFS. For NIPP, HOL had 2.28 inseminations while SRB x HOL and BS x HOL were on a level of 1.76 and 1.77 inseminations, respectively. The results for SRB x HOL crosses in general agree well with the results by Heins et al. (2006abc).
Crossbreeding under European conditions
Concluding from the above, it appears that crossbreeding indeed has distinct advantages for functional traits such as survival and fertility. However, all results mentioned with the exception of the special case of New Zealand, were obtained from comparisons of F1-animals with purebreds. However, for dairy breeders and producers who normally intend to breed their own replacements, an important question is how to go on from F1-animals. Backcrossing to Holstein will reduce heterosis effects and thus may not be advisable. Rotational crosses would require sophisticated breeding strategies within each herd, and up to now no results exist from experiments under European or North American conditions.The distinct advantages of F1-animals can only be fully exploited in a hierarchical system with specialized breeding and production tiers. Commercial milk producers would then buy F1-animals and replace them regularly, concentrating on producing milk. Under present conditions, there is no way for breeding herds to produce their own replacements and F1-animals for sale. This could only be envisaged if semen sexing would become common practice. Up to now, only a limited amount of sexed semen is available at high price and low expected pregnancy rates, but it may find increasing acceptance, especially for inseminating virgin heifers in well managed herds, until further refinements of the procedure of sexing semen are achieved. In view of the current demand and attractive price for purebred Holstein breeding stock, breeders are more likely to stick to the Holstein breed as first choice for well managed herds.
The indication of specific combining ability of individual bulls will not surprise plant or animal breeders. But it should be taken as a note of caution against recommending crossbreeding as an easy solution to current problems. Systematic crossbreeding would require a long-term breeding plan to progenytest young bulls for purebred and crossbred performance. In contrast to chickens or pigs, the information from sisters will be limited due to low reproductive rates. Small breeds like SRB would need to organize crossline testing of all their young bulls on a sufficient scale – in addition to their current pureline testing – with subsequent selection for combining ability.
Conclusions
Crossbreeding in dairy cattle under the conditions as found in Europe and North America is a tempting alternative. F1-animals show distinct advantages with respect to fertility and survival while their production characteristics reach those of the Holstein breed. However, it is not clear how individual cattle holders should continue from the F1-animals since producers like to breed their own replacements. The use of F1-animals and a separation of the breeding and the commercial tier would be facilitated if sexed semen could routinely be used for cows at all ages which is not the case at present. Data from long-term experiments of rotational crosses are not available except for the specific case of New Zealand.HEINS, B. J., L. B. HANSEN, and A. J. SEYKORA (2006a): Production of pure Holsteins versus crossbreds of Holstein with Normande, Montbeliarde, and Scandinavian Red. J. Dairy Sci. 89: 2799–2804.
HEINS, B. J., L. B. HANSEN, and A. J. SEYKORA (2006b): Calving difficulty and stillbirths of pure Holsteins versus crossbreds of Holstein with Normande, Montbeliarde, and Scandinavian Red. J. Dairy Sci. 89: 2805–2810.
HEINS, B. J., L. B. HANSEN, and A. J. SEYKORA (2006c): Fertility and survival of pure Holsteins versus crossbreds of Holstein with Normande, Montbeliarde, and Scandinavian Red. J. Dairy Sci. 89: 4944–4951.
LIC (2006): Dairy Statistics 2005-2006. www.lic.co.nz.
LOPEZ-Villalobos, N., D. J. GARRICK, H. T. BLAIR , and C. W. HOLMES (2000a): Possible effects of 25 years of selection and crossbreeding on the genetic merit and productivity of New Zealand dairy cattle. J. Dairy Sci. 83:154–163.
LOPEZ-Villalobos, N., D. J. GARRICK, C. W. HOLMES, H. T. BLAIR, and R. J. SPELMAN (2000b): Effects of selection and crossbreeding strategies on industry profit in the New Zealand dairy industry. J. Dairy Sci. 83:164–172.
LOPEZ-Villalobos, N., D. J. GARRICK, C. W. HOLMES, H. T. BLAIR, and R. J. SPELMAN (2000c): Profitabilities of some mating systems for dairy herds in New Zealand. J. Dairy Sci. 83: 144-153.
MCALLISTER, A. J., A. J. LEE, T. R. BATRA, and C. Y. LIN (1994): The Influence of additive and nonadditive gene action on lifetime yields and profitability of dairy cattle. J. Dairy Sci. 77: 2400-2414.
TOUCHBERRY, R. W. (1992): Crossbreeding effects in dairy cattle: the Illinois experiment, 1949 to 1969. J. Dairy Sci. 75: 640-667.
VAN RADEN, P. M. and A. H. SANDERS (2003): Economic merit of crossbred and purebred US dairy cattle. J. Dairy Sci. 86: 1036-1044.